Aspects hémodynamiques
Véritable carrefour veineux, le creux poplité voit l’abouchement de trois plans veineux superposés : la veine saphène externe et ses collatérales qui drainent la zone hypodermique, les veines jumelles interne et externe qui drainent la zone intramusculaire et, enfin, la veine poplitée, parfois doublée d’un canal collatéral, qui collecte le sang de la zone intermusculaire (à partir des veines péronières, tibiales antérieures et postérieures).
D’un point de vue fonctionnel, à l’état physiologique, les veines jumelles intramusculaires représentent un réservoir veineux placé au centre du « cceur périphérique » que constitue la pompe musculaire du mollet. Lors de la contraction musculaire, le sang est chassé à très fort débit vers la veine poplitée. Lors du relâchement musculaire ou en cas d’à-coups de pression dans la veine poplitée (par exemple, lors d’un effort en apnée), la portion extramusculaire des veines jumelles qui chemine dans le tissu cellulaire lâche du creux poplité va se dilater et amortir les à-coups de pression.
Lorsque les veines jumelles sont anormalement dilatées (figure 75A), elles forment un réservoir sanguin plus important. En cas de contraction musculaire brusque associée à un effort en apnée, le sang des veines jumelles intramusculaires est éjecté à fort débit vers le réseau poplité (figure 75B). L’hyperpression veineuse profonde (effort en apnée, obstacle à l’évacuation du sang par le réseau veineux profond) rend difficile le drainage. Il s’ensuit une élévation importante de la pression poplitée. Dans ce cas de figure, la saphène externe représente un moyen d’évacuation du sang en excès des veines jumelles et constitue ainsi un deuxième « amortisseur ».
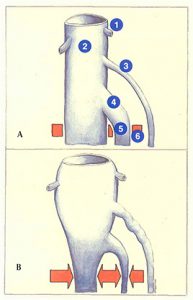
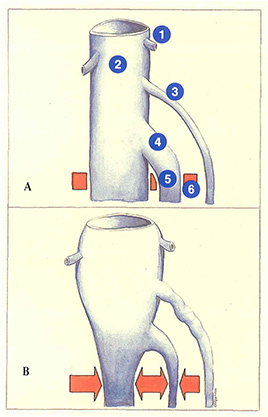
On comprend dès lors qu’une crossectomie saphène externe associée à un éveinage va éliminer ce mécanisme d’amortissement de l’hyperpression et favoriser l’apparition à court terme d’une perforante au creux poplité.
De ces éléments peuvent être tirées deux conclusions :
- une ectasie importante des veines jumelles internes peut compromettre les résultats de la chirurgie de la saphène externe en favorisant une récidive précoce par des perforantes du creux poplité;
- une ligature du tronc commun des veines jumelles sera sans conséquence quand sa terminaison s’effectue en lambda ou quand le réseau veineux jumeau interne peut se drainer par une autre voie dans le réseau poplité. En revanche, en l’absence d’une telle solution de drainage, cette ligature entraînera une hyperpression dans le réseau intramusculaire lors de la contraction et sera à l’origine de reflux vers les perforantes jumelles. A ce titre, plusieurs auteurs ont déconseillé la ligature du tronc commun des veines jumelles au creux poplité.
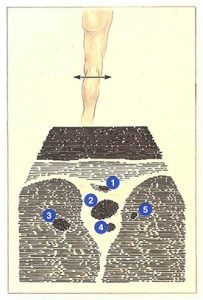
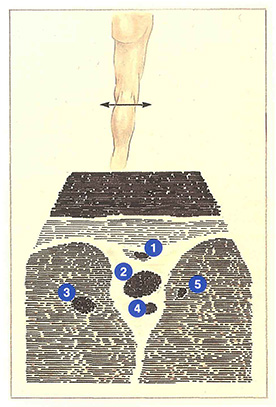
Sur l’écran d’échographie (figure 76), les veines du creux poplité ont une disposition caractéristique. Placée entre les masses musculaires et au-dessus de l’artère poplitée, la veine poplitée a le calibre le plus important. Dans 20 % des cas, un canal veineux collatéral double la veine poplitée. De même diamètre que l’artère poplitée, il s’en distingue par son caractère compressible par la sonde d’échographie.
Au-dessus de la veine poplitée, la saphène externe chemine dans un dédoublement aponévrotique. Les veines jumelles apparaissent dans les masses musculaires qui encadrent la veine poplitée.
Pour en savoir plus
Daniel C. Le syndrome des veines jumelles. Actualité Vasculaire Internationale, 1992; 6 : 16-22.
Gillot C. La perforante polaire inférieure du muscle jumeau interne. Phlébologie, Édition Médicale Média Internationale, Congrès de Bruxelles, vol 1, 1983.
Thierry L. Physiology of the muscular veins. Phlébologie, Éditions John Libbey Eurotext, Montréal 1992.
Van der Stricht J. Staelens I. Les veines musculaires du mollet. Phlébologie 1994; 47, 2: 135-43.